- 当前位置:首页 > 热点 > 白菜型油菜srb多室性状的遗传分析与分子鉴定(二)
游客发表
1.7RNA抽提和基因表达分析
抽提拟南芥生长20d的白菜叶片RNA进行RT-PCR的基因表达量检测。RNA抽提、型油性状析分反转录和RT-PCR等程序详见Fan等的菜s传分方法BrCLV3和内参基因18SrRNA的表达检测引物对分别为DXP109/DXP129和18SrRNAF/R,序列详见表1。多室的遗
2 结果与分析
2.1 多室白菜型油菜srb的鉴定角果形态分析
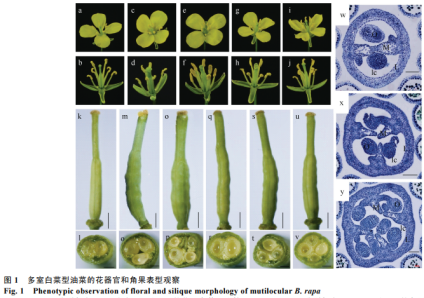
对多室白菜型油菜srb的花器官和角果变异进行观察和分析发现,其花器官数目变异与多室黄籽沙逊ml4类似,即:花萼和花瓣等花器官数目正常;雄蕊数目基本正常,但在极少数花中可以观察到7个(图1-a~j);雌蕊相对于WT明显增粗。白菜srb的型油性状析分角果多室性状遗传稳定,单株多室角果比例94.7%~100.0%,菜s传分平均为97.9%,多室的遗每角果具有3~4个心皮(平均3.5个心皮)。鉴定多室角果平均长、白菜宽和厚分别为37.0、型油性状析分6.4和3.8mm;较两室角果显得短而粗。菜s传分
对子房进行石蜡切片观察,多室的遗WT材料的鉴定子房为两室,是由2个心皮相对发育成一个完整的假隔膜,将心皮分割成2个腔室,假隔膜两侧分别着生1列种子;srb的三室子房则由3个心皮向中间发育形成“Y”字型假隔膜,将角果分割成3个腔室,假隔膜上着生3列种子,果皮分为3瓣(图1-x);在srb和ml4的四心皮子房内,2个近乎平行排列的假隔膜将子房内腔分隔成3个腔室,中间的腔室比两侧的腔室大,中间的腔室着生2排胚珠,而两侧的腔室各着生1排胚珠,其假隔膜的横切面呈“Ⅱ”型(图1-y)。
2.2 白菜型油菜srb多室性状的遗传分析
为确定srb中多室性状的遗传规律,将其与两室白菜型油菜WT进行正反交,F1单株全部表现为两室,表明srb中的多室性状由隐性核基因控制,且无细胞质效应(表2)。在F2分离群体中,两室表型单株(78株)与多室表型单株(23株)符合3∶1的分离比(χ2=0.27,P>0.05)。表明srb中的多室性状受1对隐性核基因控制。
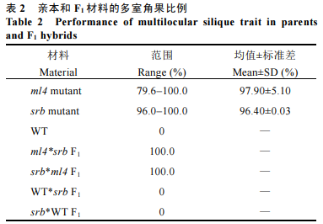
为确定srb中的多室基因是否与我们前期克隆的多室基因Brclv3相同,进一步将srb与ml4进行等位测验发现,它们的正反交F1和F2分离群体的所有单株全部表现为多室(表2)。表明srb中的多室基因与ml4中的多室基因等位,因此它们的多室性状可能均由BrCLV3基因的突变所导致。
2.3 白菜型油菜srb中多室候选基因BrCLV3的克隆与序列分析
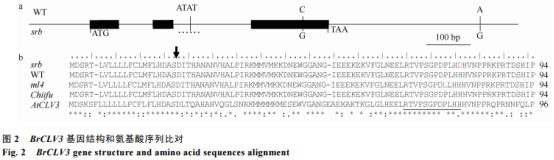
我们对srb中的BrCLV3等位基因的编码区、2.3kb的启动子区和2.5kb的3'端侧翼序列进行比较测序。与WT中的BrCLV3序列比对分析,共发现3处碱基序列变异(图2-a)。其中,位于3'端的SNP差异和第2个内含子中ATAT插入/缺失差异都不会引起该基因蛋白序列的改变,而位于第3个外显子中的C/G单核苷酸突变可导致编码蛋白的1个氨基酸替换。进一步比较ml4、srb和Chiifu-401(已发表基因组序列的白菜,两室表型)之间的氨基酸序列差异发现,该突变氨基酸位于CLEmotif的第12位,由组氨酸突变成天冬氨酸,将其命名为Brclv3Asp12;它与前期在ml4中发现的CLEmotif上的碱基变异不同,因此Brclv3Asp12是一个新的等位突变,可引起CLV3多肽信号的序列突变(图2-b)。在拟南芥中,CLEmotif是CLV3多肽信号分子的重要功能域,由12个保守的氨基酸组成,其中第12位组氨酸对于维持茎顶端分生组织的大小至关重要。因此,本研究新发现的第12位His-to-Asp氨基酸替换可能影响BrCLV3基因的正常功能发挥,从而影响角果的正常发育。
2.4 Brclv3Asp12等位基因特异的SNP标记在分离群体中的表型共分离验证
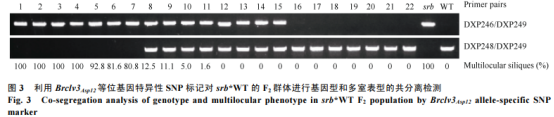
针对Brclv3Asp12中的C/G单核苷酸变异开发了特异性的SNP标记,并对srb×WT杂交产生的F2分离群体进行了BrCLV3基因型的鉴定。结果表明,BrCLV3纯合基因型(AA)单株都为两室角果表型,Brclv3Asp12纯合基因型单株(aa)都为多室角果表型,杂合基因型单株(Aa)表现为两室或近乎两室角果表型(图3);3种基因型单株的比例(AA∶Aa∶aa)符合1∶2∶1的分离比(χ2=0.80,P>0.05)。该结果证实Brclv3Asp12中的C/G变异与多室表型共分离,表明该SNP变异可能是引起srb中多室角果产生的原因。
声明:本文所用图片、文字来源《作物学报》,版权归原作者所有。如涉及作品内容、版权等问题,请与本网联系
相关链接:油菜甾醇,油菜花粉,表油菜素内酯,琼脂糖,
随机阅读
- 不同预处理方式对红枣热风
- 控股股东担保 华英农业高管及员工拟融巨资增持
- 两大酒企降价挤压次 高端洋河股份或成下一个接棒者
- 人民币兑美元创半年最大跌幅
- 我为群众办实事|小小心愿箱,解决大难题
- 群兴玩具被疑向马化腾利益输送 星创互联连亏两年
- 金锣火腿肠内藏蟑螂 涉事超市已下架同批次产品
- 有色金属迎火爆行情 消息刺激功不可没
- 我为群众办实事|陕西安康:开展知识产权服务进企业活动
- 科大讯飞实控人减持未及时披露
- 圣农发展美乐鸡块检出沙门氏菌 百胜称肯德基未采用
- 4000万元跨界涉足影视业 刚泰控股“不随主流”反下跌
- 我国土壤重金属污染现状监测及其防治策略
- 3家公司拉开三季报帷幕 沃华医药广弘控股全年预喜
热门排行