- 当前位置:首页 > 探索 > 非生物胁迫对橡胶树热激转录因子家族成员表达的影响(二)
游客发表
2.2 干旱对热激转录因子基因表达的非生影响
在橡胶树30个Hsfs家族成员中,有28个成员响应干旱胁迫,物胁只有HbHsfA1c和HbHsfA5b两个基因不响应干旱胁迫(图2)。迫对在干旱处理的橡胶响橡胶树‘93-114’叶片中,HbHsfA3b、树热HbHsfA4a、激转家族HbHsfA4c、录因HbHsfA4d、表达HbHsfA5a、非生HbHsfA6a、物胁HbHsfA7a、迫对HbHsfA9a、橡胶响HbHsfA9b、树热HbHsfB1a、激转家族HbHsfB3a、录因HbHsfB4a和HbHsfB4b共13个基因响应干旱处理并呈持续显著上调表达模式,其中HbHsfA7a、HbHsfA7a和HbHsfB4b的上调幅度较大,在干旱处理4h时它们的上调幅度是0h的3倍以上,对干旱胁迫比较敏感。HbHsfA1b、HbHsfA2b、HbHsfA3a和HbHsfA4b响应干旱胁迫显著下调表达,其中HbHsfA1b和HbHsfA3a在干旱处理中后期持续显著下调表达,而HbHsfA2b和HbHsfA4b分别在干旱处理2h和24h时极显著下调表达,响应干旱胁迫的时间有明显差异。HbHsfA1a、HbHsfA2a、HbHsfB2c、HbHsfB3b、HbHsfB4d、HbHsfC1a和HbHsfC1b在干旱处理的前期上调表达,在后期下调表达;而HbHsfA8a和HbHsfB2b的表达模式与之相反,它们在干旱处理的前期下调表达,在后期上调表达(图2)。HbHsfB2a在干旱胁迫下先显著上调表达,然后显著下调,后期又极显著上调表达;而HbHsfB4c在干旱胁迫下先显著下调表达,然后显著上调,中后期又极显著下调表达(图2)。
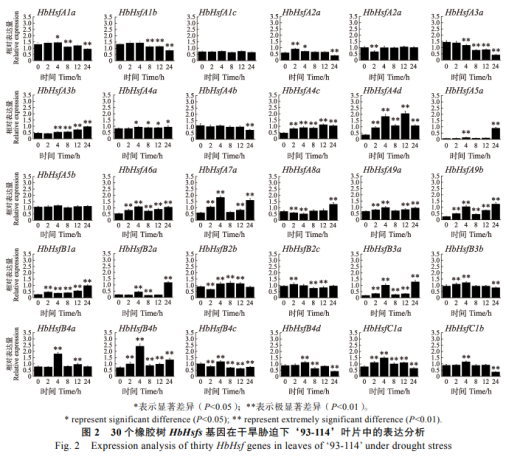
2.3 高盐(NaCl)对热激转录因子基因表达的影响
在橡胶树30个Hsfs家族成员中,有29个成员响应NaCl高盐胁迫,只有HbHsfA1c不响应高盐胁迫(图3)。在高盐处理的橡胶树‘93-114’叶片中,HbHsfA2b、HbHsfA3b、HbHsfA4a、HbHsfA4b、HbHsfA5b、HbHsfA7a、HbHsfB1a、HbHsfB2a、HbHsfB2b、HbHsfB2c、HbHsfB3a、HbHsfB3b、HbHsfC1a和HbHsfC1b共14个基因响应高盐胁迫并呈持续显著上调表达模式,尤其HbHsfB2a和HbHsfB3a响应高盐胁迫上调幅度最大,在高盐处理2h时它们的转录水平是0h的2倍以上,对高盐胁迫比较敏感,而HbHsfA2b对高盐胁迫的反应比较滞后,在12h之前无响应,在处理24h时HbHsfA2b的表达迅速上调,几乎是0h的3倍。HbHsfA1a、HbHsfA1b、HbHsfA3a和HbHsfA4c响应高盐胁迫显著下调表达,其中HbHsfA1a、HbHsfA3a和HbHsfA4c在高盐处理中后期持续显著下调表达,而HbHsfA1b在高盐处理8h时极显著下调表达。HbHsfA2a、HbHsfA4d、HbHsfA5a、HbHsfA9b、HbHsfB4a、HbHsfB4b和HbHsfB4d、在高盐处理的前期显著上调表达,在后期显著下调表达,与这些基因表达模式相反的基因HbHsfA8a和HbHsfB4c分别在高盐处理的前、中期显著下调表达,在中、后期显著上调表达。HbHsfA6a和HbHsfA9a在高盐胁迫前期先显著上调表达,然后显著下调,后期又显著上调表达(图3)。

3 讨论
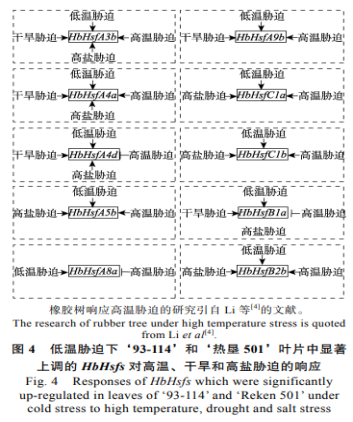
本研究分析了30个橡胶树HbHsf家族成员在低温胁迫下橡胶树‘93-114’和‘热垦501’叶片中的表达模式。结果表明,在HbHsf家族中有10个基因不仅在抗寒橡胶树种质‘93-114’叶片中上调表达,也在不抗寒橡胶树种质‘热垦501’叶片中显著上调表达。基因表达在不同组织中既有组织的特异性也有组织的相似性。比如上述叶片中上调表达的10个基因中,在低温条件下,HbHsfA3b、HbHsfA4a、HbHsfA5b、HbHsfA8a、HbHsfC1a、HbHsfC1b和HbHsfB2b在橡胶树‘93-114’和‘热垦501’叶片中均呈显著上调表达模式,同时也在相应橡胶树的树皮中呈上调表达的模式,呈现组织的相似性。但是其他3个基因在树皮和叶片中的表达模式不一致,呈现组织的特异性。在这2个不同耐寒性的橡胶树种质中都响应低温胁迫,但是响应程度不同。尤其HbHsfA4a在低温胁迫下橡胶树‘93-114’叶片和树皮中的表达量均显著高于‘热垦501’。这些基因表达量的高低可能与橡胶树抗寒性的强弱相对应。
枇杷EjHsf1(A4)响应低温诱导,通过转录调控EjHsp基因参与提高枇杷抵抗低温胁迫能力。水稻OsHsfA4d受低温胁迫后上调表达。OsHsfA4s在高盐胁迫后表达量上调。草莓FvHsfA4a受低温和干旱胁迫上调表达。橡胶树HbHsfA4a和HbHsfA4d是EjHsf1、OsHsfA4和FvHsfA4a的同源基因,同样受低温、干旱和高盐胁迫后表达量上调表达(图4),另外,HbHsfA4a受高温胁迫上调表达,而HbHsfA4d受高温诱导下调表达。水稻OsHsfA3和OsHsfA9受低温胁迫后上调表达。草莓FvHsfA3a在高温、低温、干旱和高盐胁迫下上调表达,FvHsfA9a在低温和高盐胁迫下上调表达。辣椒CaHsfA3/A9受高盐胁迫上调表达。同源基因HbHsfA3b在高温、低温、干旱和高盐胁迫下的橡胶树‘93-114’叶片中上调表达,HbHsfA9b响应高温、低温和干旱上调表达(图4)。小麦B2亚家族TaHsf3响应高温、低温、干旱和高盐的诱导,过表达该基因激活下游基因热激蛋白HSP70,从而提高拟南芥植株的耐热性和抗寒性。鹰嘴豆CarHsfB2在高温、高盐和干旱胁迫下上调表达,过表达CarHsfB2能够提高拟南芥植株的耐热性和抗旱性。但是OsHsfB2b对水稻的抗旱性起负调控作用。B1亚家族基因响应高温、干旱和高盐的诱导。同源基因HbHsfB2b在高温、低温和高盐胁迫下的橡胶树‘93-114’叶片中上调表达,HbHsfB1a响应高温、低温、高盐和干旱上调表达(图4)。水稻OsHsfC1a/b、OsHsfC2a/b在低温胁迫下表达量显著上调,其中OsHsfC1对低温胁迫最敏感。拟南芥的AtHsfC1同样可被低温诱导,表达量极显著上调。另外,在葡萄、胡萝卜和甘蓝等园艺植物中也发现VaHsfC1a、DcHsfC16和BraHsfC039受低温胁迫表达量显著上调。由此可见,C亚家族成员在植物抵抗低温胁迫中也起着重要的作用。此外,小麦TaHsfC2a受高温胁迫上调表达,过表达TaHsfC2a可上调干旱和高温诱导基因的表达,提高转基因植株的耐热性。OsHsfC1b在水稻抵抗高盐胁迫中起着重要的作用。由此可见,橡胶树HbHsfA3b、HbHsfA4a、HbHsfA9b、HbHsfC1a、HbHsfC1b和HbHsfB2b等基因响应多种逆境交叉胁迫,暗示着上述基因功能的多样性和复杂性。
高温、低温、高盐、干旱等逆境常常会导致植物体内发生氧化胁迫。Hsfs是上调ROS清除酶的活性和HSP基因表达的关键因子,利于提高植物对逆境胁迫的抗性。拟南芥体内的AtHsfA3受活性氧诱导表达量上调,过表达AtHsfA3的拟南芥植株中,该基因可直接结合肌醇半乳糖苷合成酶基因GolS1和GolS2的启动子从而激活GolS1和GolS2的转录,肌醇半乳糖苷的含量显著增加,利于清除羟基自由基,以免植物细胞受到逆境胁迫的伤害。过表达CmHsfA4能够上调转基因菊花中CmHSP70、CmHSP90等基因的表达,提高SOD、APX、CAT等活性氧清除酶的活性,降低植物细胞内活性氧的含量,从而增强了转基因植株的抗逆性。东南景天SaHsfA4c转基因拟南芥可激活ROS清除酶的活性和上调HSP的表达,降低活性氧的积累,从而增加植物对逆境胁迫的抗性。水稻体内OsHsfC2a响应H2O2诱导表达量上调,可以提高水稻对氧化胁迫的敏感性和抵抗能力。由此可见,橡胶树HbHsfA3b、HbHsfA4a以及HbHsfC亚家族成员可能参与氧化胁迫反应,保护细胞免受氧化损伤,提高橡胶树的抗逆性。
声明:本文所用图片、文字来源《热带植物学报》,版权归原作者所有。如涉及作品内容、版权等问题,请与本网联系
相关链接:胡萝卜,辣椒,半乳糖苷合成酶
随机阅读
热门排行